
垃圾DNA的来源——染色体不对称分配
微管的不对称修饰,可以解释染色体的优先遗传机制。
最近的研究表明,两条姐妹染色体的着丝粒(减数分裂过程中介导染色体分离的、与微管结合的染色体区域)在雌性减数分裂过程中相互竞争以获得遗传。鉴于此,介导准确染色体分离的基本DNA序列实际上是侵入我们基因组的“自私”(或寄生)遗传元件。对此,Akera等人提供了迄今为止最详细的分子机制,他们解释了寄生DNA序列如何利用卵母细胞减数分裂的不对称性来确保自身的特征能够遗传下去,并因此在种群中得以传播的机制。
着丝粒DNA由包含1000个拷贝的、非常短的(100-300个碱基对)的一个序列组成,这个序列在拷贝数和序列上快速增加。科学家们对此有两种完全不同的猜想。一种猜想是,这些DNA可能具有重要的功能;另一种猜想则是这些短的DNA序列可能是自私的,只会促进其自身的遗传,而对宿主生物体没有任何功能益处。破译这个谜题非常重要——着丝粒重复序列是我们基因组中最丰富的非编码DNA,但到目前为止,我们却不知道它们是否有作用,如果有,作用是什么。最近的一些研究结果显示,着丝粒DNA可能确实是自私DNA,也就是对生物体自身无任何功能益处的垃圾DNA。
标准的实验室小鼠品系有20个不同的染色体,每个染色体的一端都有着丝粒(端着丝粒,telocentric)。相反,某些隔离的野生小鼠群体具有10条染色体,每条染色体由两个端着丝粒染色体融合成一个染色体而成,着丝粒位于中间(中央着丝粒,metacentric)。实验室小鼠和野生小鼠杂交后得到的雌性后代表现出一种名为减数分裂驱动的特性。这些雌性小鼠的卵母细胞在减数分裂时,并不会随机地将端着丝粒染色体或中央着丝粒染色体遗传给卵子,而是有倾向性地进行遗传。目前人们对这些发现的了解还不深入,因为这一现象只解释了为什么野生种群的老鼠往往携带的都是端着丝粒染色体,或者都是中央着丝粒染色体,但没有解释具体机制。
最近的研究表明,相对于不经常被遗传下来的染色体,优先传播给后代的染色体的着丝粒重复序列的拷贝数更高,可达前者的6倍,并且携带的动粒蛋白(kinetochore protein)更多。具有更多重复拷贝和更多动粒蛋白的优先遗传的着丝粒被称为“强”着丝粒,并且它们优先朝纺锤体的卵子方向移动。具有较少重复拷贝的“弱”着丝粒则优先地朝质膜移动,在染色体分离后它们将被保留在极体中,并随着极体的消失而消失。即使在具有相同染色体数目的动物杂交得到的后代(即姐妹染色体都具有端着丝粒或都具有中央着丝粒)中,着丝粒重复序列的数目也会影响分离偏好。这表明减数分裂驱动发生在每个杂交品系中,而不只发生于具有不同染色体数目的动物杂交的罕见情况下。
简单地增强一个着丝粒,并不足以解释这种优先遗传现象。动粒是位于着丝粒上两侧的蛋白质结构。在细胞分裂过程中,纺锤体的纺锤丝需连接到染色体的动粒上,牵拉染色体到细胞两极。每个动粒与微管的末端结合,产生力来牵引染色体向纺锤体的一极移动。如果一对着丝粒产生的作用力较大,那么在染色体分离之前,这对染色体就已经开始偏向纺锤体的一极,而这会导致一对染色体的两个着丝粒与纺锤体同一侧发出的微管结合。如果在雌性减数分裂过程中,这一错误没有得到纠正,那么最后得到的卵子就会多一条染色体,受精发育后形成的后代往往会流产或早逝(如发生二十一三体综合征)。
Akera等人进一步解释了染色体不对称分配的机制。纺锤体首先在卵母细胞中心附近组装,这里的纺锤体在结构上是对称的,强和弱的一对着丝粒各自随机地向两个纺锤体极移动。纺锤体微管向质膜(极体一侧)的迁移使结合的染色体靠近质膜。来自染色体的近距离信号[鸟苷三磷酸(GTP)-Ras相关核蛋白(RAN)]局部激活质膜上的邻近依赖性信号[GTP-细胞分裂调控蛋白42同系物(CDC42)]。随后,GTP-CDC42使得靠近质膜的微管比远离质膜的微管更倾向于酪氨酸化。动粒与重度酪氨酸化的微管的结合相对不稳定,而微管与强动粒的结合也相对不稳定。因此,质膜近端的酪氨酸化微管与强动粒的结合相对不稳定,容易中途断裂,导致动粒对方向发生翻转,强动粒开始与卵子一侧的微管结合,而弱动粒与质膜一侧的微管结合。接着,强动粒附着的染色体会进入卵子,并在胚胎中得到遗传(图:雌性减数分裂过程中的优先遗传)。
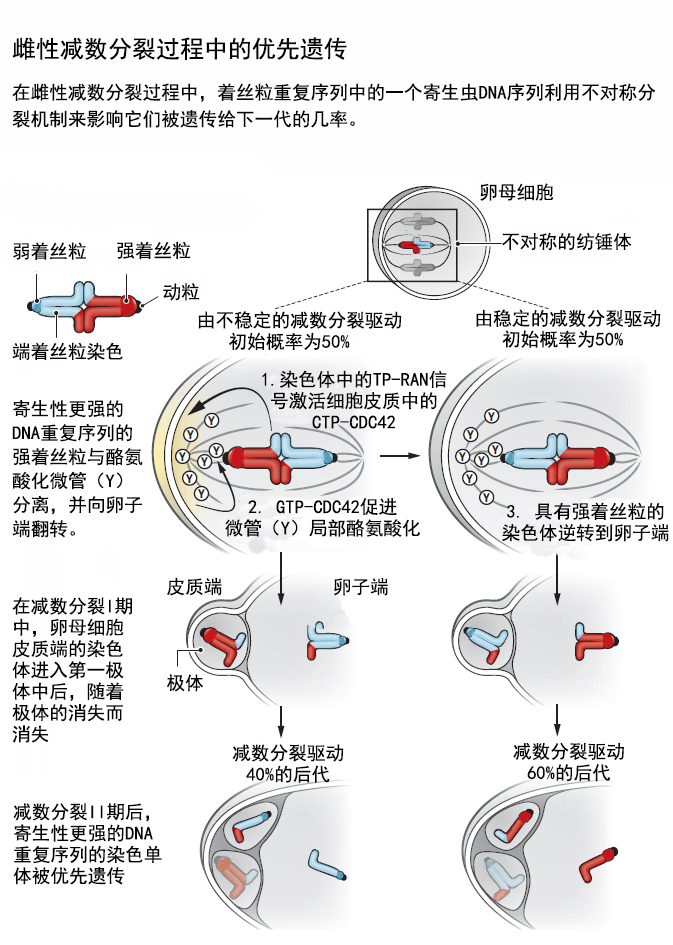
这个模型引起人们的极大兴趣,因为它体现了目前成对的动粒如何与相反两极的微管正确地结合的机制。这种结合事件完全是随机的,不正确的结合容易断裂,而正确的结合则是稳定的。未来我们需要进一步了解这个过程。首先,GTP-CDC42局部增加微管蛋白酪氨酸化(或抑制去酪氨酸化)的机制仍有待研究。第二,多大程度的酪氨酸化的微管会与动粒脱离,或者为什么一个强的动粒会更易与微管脱离。酪氨酸化的微管蛋白是细胞骨架相关蛋白富含甘氨酸(CAP-Gly)结构域的蛋白质的首选结合位点,而最近的研究表明,马达蛋白、胞质动力蛋白优先运输酪氨酸化微管。因此,进一步研究微管的翻译后修饰机制将有助我们深入挖掘自私DNA如何影响我们的基因组。
原文检索:
Francis J. McNally. (2017) Competing chromosomes explain junk DNA. Science, 358: 594-595. 张洁/编译



{kind=link}